ベッコウトンボが減少した原因についての一考察 倉品治男 H.Kurasina: A study for the decreasing cause ofLibellula angelina |
||
| はじめに | ||
| 図1の左の図は1969年におけるベッコウトンボの生息状況 (環境庁,1993)を示し、右は2000年における生息状況 (日本蜻蛉学会 ベッコウトンボ部会,2000)である。 | ||
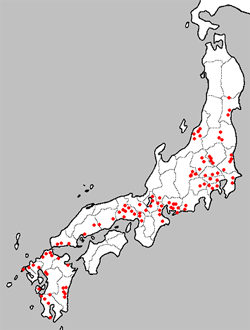 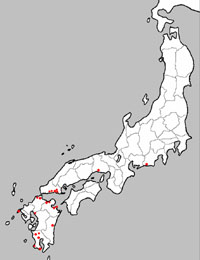 図1 ベッコウトンボ生息状況 1969年(左)と2000年(右) |
||
|
1969年には宮城県北部から鹿児島県まで、主として本州を中心に分布していたが、2000年の本州における生息地は3ヶ所しかなく、九州においても大分,鹿児島を除けば絶滅または絶滅寸前の状態となっている。 1 絶滅の第一ステージ |
||
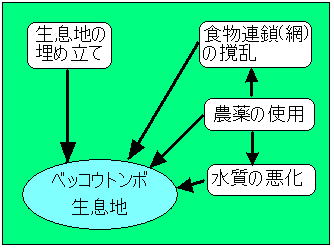 図 2 絶滅の第一ステージ |
||
| その為に、止水域に生息する生物種の中で大型注水植物が繁茂する浅い水域に生息する生物種群が、埋め立てによる生息環境の消滅という打撃をより強く受ける結果となった。(図2) あわせて、1950年代後半から本格的に普及した農薬の直接的・間接的影響も否定出来ない。 農薬、特に各種の殺虫剤の使用によって農耕地周辺の生物相は壊滅的な撹乱を受け、捕食動物においては直接的な影響だけでなく、餌動物の減少や消滅といった食物連鎖(網)の撹乱による間接的影響も強く受ける結果となった。(図2参照) 一口に農薬といっても、殺虫・殺菌・除草さらに化学肥料も加えるとすれば、使用されている薬品類の数はきわめて多く、どのような農薬がどのように作用したかについては一般には何も解っていないのが実情である。 しかし、埋め立てや農薬の影響による負荷はベッコウトンボだけに選択的に加わったわけではなく、同一環境条件下に生息する全ての昆虫類に均等に加わったはずであり、ベッコウトンボだけが減少した説明としては不十分である。 筆者は1997年の本誌6月号で、生息池から半径2km以内にあって、相観上ベッコウトンボ生息地としての条件を充たしている池には必ず当該種が飛来することを記述した。 その後、飛来が見られても「ベッコウトンボが生息する池」と「生息しない池」があることに着目して現状を把握すべく調査を進めた。 その結果、「飛来はしても生息が認められない池」では農業排水や家庭からの排水が流入し、「生息が認められる池」ではそれらの流入が無いか、あっても極わずかであるという、特徴ある現象を知見することが出来た。 しかし、どちらのタイプの池にもベニイトトンボ、コバネアオイトトンボ、トラフトンボ、アオヤンマ、ヨツボシトンボ、マイコアカネなどのベッコウトンボ生息池に特有のトンボ相が見られることは、知見した限りでは注目すべき現象である。 もちろん、農薬の流入量が増大し、直接的・間接的な影響量が増えれば、これらの種も順次姿を消すのはいうまでも無いことであるが、まず始めに姿を消すのがベッコウトンボなのである。 これらのことから、ベッコウトンボは農薬等に使用されている何らかの薬物に過敏な種ではないかとの疑いが持たれる。 もう一つの可能性としてはベッコウトンボの生態的な特徴に起因するのではないかという疑いである。 ベッコウトンボの幼虫は他のトンボ類の幼虫と比較して採餌のための徘徊行動が少ない事が多くのトンボ研究者によって指摘されている。 この性質は餌となる動物の生息密度が高ければ同種間での共食いを防ぎ、動き回る無駄なエネルギーを少なくする最高の方法である。 しかし、餌の密度が低くなれば生存できる可能性は低くなり他のトンボ類に先がけて消滅する事になる。 除草剤の止水環境への流入が直接或は間接に動物プランクトンやフサカの幼虫を減少させるとすれば、餌動物の密度が低下する事になりベッコウトンボ幼虫の生存率は一気に低下するであろう。 しかし、現在の段階ではその影響が直接的なものなのか、間接的なものなのか決定する決め手がなく、両方の可能性を視野に入れながら調査を継続中である。 2 絶滅の第二ステージ 前掲の通り、各地のベッコウトンボ生息地は次々と消滅したが、埋め立てからも、農薬の影響からも免れた生息地は各地に孤島状に残った。 次に、この孤島状に残った生息地に何が起こったのかについて考えてみたい。 |
||
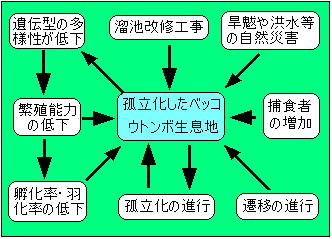 図3 孤立した生息地における絶滅のプロセス |
||
| 孤立した生息地は図3に示したような旱魃などの自然災害や池内植物の遷移に伴う陸化、オオクチバス・ウシガエル・コイ・アメリカザリガニ等を池内に持ち込むことによる新たな捕食者の出現、そして最も決定的なダメージを及ぼす溜池の改修工事等の影響を受けることによって消滅へと向かうことはいうまでもない。 しかし、ベッコウトンボ個体群への明らかな負荷は認められないにもかかわらず次第に減少し消滅する例も多く観察されている事に着目し、以下にそのような例の絶滅要因について考察を試みた。 ベッコウトンボは羽化後ほとんどその水域を離れることなく未熟期を過ごし、成熟後羽化水域に戻ると言われており、(福井,1988)この習性がベッコウトンボ個体群の行動を特徴付ける一つとなっている。 以下、この習性を「生息地に留まる性質」と呼ぶこととし、その「生息地に留まる性質」についての知見をまとめた。 ベッコウトンボは強い「生息地に留まる性質」を示すといわれているが、その一方で個体群から移出する行動も知られている。 これらの行動を把握するために各地で標識調査が行われたが、いずれの調査においても移出個体数が把握できなかったことから非移出個体数も明らかにすることが出来ず、「ほとんどが生息地に留まる」という表現にならざるを得なかった。 筆者は1997年に大分県中津市野依新池において標識調査を行った。 再捕獲結果に雌雄の性比、確認率を勘案した結果 移出性については5%〜10%と推測し、 従って、「生息地に留まる性質」については90%〜95% と推測した。 この結果からも確かに高い「生息地に留まる性質」が立証できた。 尚、前生殖期においては発生水域周辺(半径約500m)の草地に移動して過ごすが、移出個体とはこの生息圏外の池で繁殖行動を行っていた個体を指す。 高い「生息地に留まる性質」は他の生息適地を探し回ることにより派生する様々な危険性を回避出来ると共に、現生息地で種の存続を図ることだけにエネルギーを費やすことが出来る優れた生存戦略である。 但し、この生存戦略は生息地が安定した自然環境下にあってこそ有効となる。 しかし、この事は、自然災害による生息地の消滅や病気の発生に対しては一気に個体群が消滅する大きなリスクを負うことになるであろう。 ベッコウトンボの移出性向はそれらのリスクヘッジに役立つと共に、個体群間の遺伝子の交流にも寄与していると推察できる。図4参照 |
||
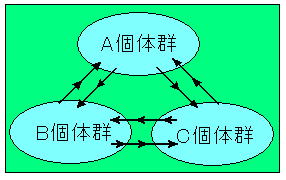 図 4 個体群間の個体の移出と移入 |
||
| しかし、孤立した生息地においては個体群の一部を移出させたとしても周辺に生息適地は既に無く、他の個体群からの移入も望めない。 言い換えれば生息地消滅のリスクを回避したり、個体群間の遺伝子の交流は望めないということである。 次に、図3に示したような負荷が加わったとは認められないにもかかわらず個体群が消滅する例として海浜部の広大な埋立地に出来た水域で突然ベッコウトンボが大発生する事例を考えてみよう。 この現象はたまたま長距離移動した一頭のメスが埋立地に相観上の生息適地を見つけて産卵した結果と思われ、こうした場合、ほぼ数年でその個体群は消滅する場合が多い。 埋立地に発生したベッコウトンボの個体群は何故消滅するのか。そのプロセスとして以下の二つのパターンを考えてみた。 その1 A.A.Berrymanはその「個体群システムの生態学(1981)」の中で、個体群システムの基本的パターンの一つとして「爆発的に膨張する個体群の変動は,変動の極めて激しい環境下に生息する個体群に見られる」と述べている。 工事終了後間もない埋立地の自然は植生遷移の初期段階であり、大きく変動する自然である。 このために、ある種の生物個体群が爆発的に膨張出来る環境下にあるといえる。 このような環境下のベッコウトンボ個体群はある環境特性下で爆発的に膨張するが、環境特性が変化するとある閾値を境に急激に消滅へと向かうと考えられる。 絶滅の第一ステージの農薬の部分で述べたように、餌を採るために動き回らないベッコウトンボの幼虫は動物性プランクトンの密度が高ければ爆発的に個体数を増やす事が出来るが、逆の場合は極端に個体数を減らしてしまう。 工事終了後間もない埋立地では動物プランクトンの密度が大きく変動すると想定され、この事がベッコウトンボ個体群の消滅にかかわっている可能性がある。 その2 視点を変えると、埋立地の個体群は孤立した個体群である場合がほとんどであり、他の個体群からの移入は期待できない。 従って、その個体群は始めに飛来した一頭のメスの直系子孫であるから、個体群内における遺伝型の多様性は低い。 このことは、ある特定条件下では高い繁殖能力を発揮する可能性を秘めているが、様々に変化する病気や寄生虫などを含めた外因に対する適応能力は低く、従って、安定した個体群にはなりえない。 おそらく、埋立地に発生した個体群には上述した二つの事が同時に起こるのであろう。 上記の事例は埋立地という環境変化の激しい生息地における個体群についてであるが、一般的にベッコウトンボの生息地は前述したように安定した環境にある。 それでは次に、孤立しているが安定した生息環境下にあるベッコウトンボ個体群がどうして消滅するのかを考察してみたい。 |
||
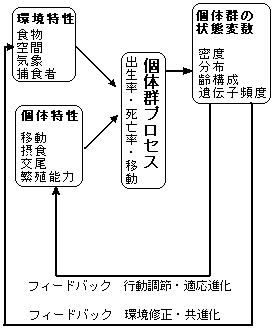 図 5 個体群システムの機構(個体群システムの生態学より) |
||
| 上述したように、ベッコウトンボ個体群の状態変数においては密度と遺伝子頻度が重要であるが、孤立した生息地であるにもかかわらず高密度で安定しているということは遺伝型の多様性も維持されていることを示唆しており、2で述べたことと矛盾してしまう。 そこで、筆者らはこの矛盾を説明するために以下の仮説を導入した。 4 亜個体群仮説の導入 |
||
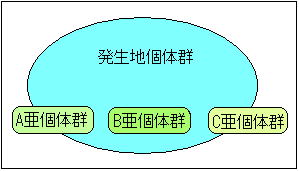 図 6 亜個体群を持つ発生地個体群の構造 |
||
| 例えば、ベッコウトンボの生息する複数の浅い池を包含する大きな湿地があったと仮定しよう。 この湿地を一つの生息地(mega)として捉えれば、その湿地内には複数の生息地個体群(sub population)があることになる。 ところが、この湿地の水位が少し上昇すれば湿地内の池は統合されて一つの池となる。 この時、この統合された池の個体群は生息地個体群とみなすことが出来るわけであるから、この生息地個体群には複数の亜生息地個体群が内包されていることになる。 上述した例は、ベッコウトンボの亜個体群の成立過程を説明したわけではなく、亜個体群を含む構造を理解してもらうための例である。 野依新池のように面積が1haしかない生息地においては、そこで発生する個体群は均一に撹拌された個体群だと思われてきた。 しかし、ベッコウトンボのように高い「生息地に留まる性質」を示す種においては池の中なら何処でもよいというのではなくごく狭い発生水域周辺に戻ってきて繁殖行動をするのではないかと考えたのである。 そして、この行動は代を重ねるうちに幾つかの集合、即ち、sub popula tion となっていったのではないだろうか、というのが筆者らの仮説である。 個体群の増減は図5に示したような環境特性や個体特性に支配されるが、一つの個体群の中に複数の亜個体群が存在すれば、生息地個体群内において、ある程度の移出入が完結し、孤立した個体群が陥る遺伝型の多様性は低下しないですむはずである。 野依新池では毎年1000個体〜3000個体の発生が1994年の調査開始以来安定して見られている。 この生息状況から判断すると、この池の個体群は高密度に安定性していて、そのシステムは負のフィードバック ループに支配されていると考えていいだろう。 野依新池や藺牟田池における個体群の安定性は安定した環境特性と亜個体群を内包することで維持される安定した遺伝特性によってもたらされているのではないだろうか、これが、筆者の推論である。 5 まとめ 筆者は上述したようにベッコウトンボが絶滅を危惧されるような状態に立ち至った原因について二つの段階に分けて推論を試みた。 第一段階の減少原因は埋め立て等による生息地の消滅と農薬などの影響であり、ベッコウトンボが農薬等に含まれるある種の薬物に対して直接的或いは、間接的に他種と比較して敏感であったことが、他種よりも際立って減少した原因であると推論した。 その結果、生息地は島状に孤立する結果となった。 第二段階において孤立した生息地の個体群は遺伝型の多様性が低下 ? 繁殖能力の低下 ? 出生率の低下 ? 遺伝型の多様性が低下というフィードバックループに支配され減衰し消滅した。 しかし、複数の亜個体群を包含している一部の生息地個体群においては遺伝型の多様性の低下は起こりにくく、環境特性に条件づけられた範囲内で安定した個体数を維持することが出来る。但し、全ての状況において図3に示したような環境要因がベッコウトンボ個体群の消滅に最も強く作用していることはいうまでもない。 おわりに 始めに述べたようにこれは調査結果に基づくものではあるがあくまでも推論や仮説に過ぎず、これらを立証するためには更なる調査と研究が必要である。 この論の弱点は孤立した生息地のベッコウトンボ個体群において遺伝型の多様性が低下するとの仮定に立脚しているが、その論拠が不十分な点にある。 また、亜個体群の存在を論じているが、羽化地点とナワバリ地点の関係を暦日に照らしての考察にすぎず、今後は羽化直後の個体をマーキングするなどの方法でさらに客観的なデータを積み重ねる必要があると思われる。 参考文献 環境庁1993. 国内希少野生動植物選定のための生息実態調査 ベッコウトンボ.55pp 浜島繁隆 「ため池の自然学入門」ため池の自然談話会編 :12-13,1994. 宝月欣二 「湖沼生物の生態学」:1998 アラン.A.ベリーマン「個体群システムの生態学」吉川 賢 訳 1985. 福井 順治 「静岡県磐田市桶ヶ谷沼におけるベッコウトンボの生態観察」:月刊むしNo 212 青木典司 「標識調査によるベッコウトンボ成虫の動態について」:昆虫と自然Vol.32 No.7 11-18 1997. 青木典司 「べっこうとんぼ発生池における個体数激減の過程と考察」 :昆虫と自然Vol.32 No.7 37-41 1997. 三時輝久 平田真二「ベッコウトンボの移動習性」昆虫と自然 Vol.32 No.727-321997 倉品治男 「ベッコウトンボの環境選択性について」昆虫と自然 Vol.32 No.7 42-45 1997. 倉品治男 「ベッコウトンボに見られる産卵を伴わない交尾に関する考察:TOMBO,XLII,2000 31-38 2000 |